https://www.cambridge.org/core/services/aop-cambridge-core/content/view/S0954422410000247
http://www.feedadditive.com/docs/S0954422410000247.pdf
Butyrate is a natural substance present in biological liquids and tissues. The present paper aims to give an update on the biological role of butyrate in mammals, when it is naturally produced by the gastrointestinal microbiota or orally ingested as a feed additive. Recent data concerning butyrate production delivery as well as absorption by the colonocytes are reported. Butyrate cannot be detected in the peripheral blood, which indicates fast metabolism in the gut wall and/or in the liver. In physiological conditions, the increase in performance in animals could be explained by the increased nutrient digestibility, the stimulation of the digestive enzyme secretions, a modification of intestinal luminal microbiota and an improvement of the epithelial integrity and defence systems. In the digestive tract, butyrate can act directly (upper gastrointestinal tract or hindgut) or indirectly (small intestine) on tissue development and repair. Direct trophic effects have been demonstrated mainly by cell proliferation studies, indicating a faster renewal of necrotic areas. Indirect actions of butyrate are believed to involve the hormono–neuro–immuno system. Butyrate has also been implicated in down-regulation of bacteria virulence, both by direct effects on virulence gene expression and by acting on cell proliferation of the host cells. In animal production, butyrate is a helpful feed additive, especially when ingested soon after birth, as it enhances performance and controls gut health disorders caused by bacterial pathogens. Such effects could be considered for new applications in human nutrition. Gastrointestinal microbial ecosystem: Feed additives: Biological role of butyrate: Trophic effects: Hormono –neuro–immuno system: Animal and human nutrition
Introduction The SCFA constitute a group of molecules that contain from one to seven carbon atoms and which exist as straightor branched-chain compounds. They mainly originate from the intestinal bacterial fermentation of plant materials such as celluloses, fibres, starches and sugars for which mammals lack the necessary enzymes to split these compounds. Among the SCFA, acetic, propionic and butyric acid are the predominant forms. Because SCFA are weak acids with a pK of # 4·8 and the pH of the gastrointestinal (GI) tract (GIT) fermentation chambers is nearly neutral, 90–99 % of the SCFA are present in the GIT as anions rather than as free acids(1). SCFA have several beneficial effects for animals. As an example, acetic and propionic acids, due to their physical and chemical properties, are used as acidifiers in pig diets in order to help overcome problems in the postweaning lag period. Moreover, dietary acidification is known to be beneficial for the performance of fattening animals(1,2). For ruminants and herbivorous animals, SCFA constitute the major substrates for energy production. Evidence also indicates that there is considerable fermentation in the caecum and colon of both human and nonherbivorous animals. Among the SCFA, butyric acid has received particular attention. It is a natural substance present in the GIT, in milk as well as in the sweat and faeces of most mammals. Butyric acid is available as the Na, K, Mg or Ca salt. The advantage of salts over free acids is that they are generally odourless and easier to handle in the feed manufacturing process owing to their solid and less volatile form. For the purpose of the present review, the term ‘butyrate’ is used interchangeably for the acid, the salt and the anion forms. In the GIT, butyrate is naturally present in high concentration in the lumen of the large intestine. It is preferentially taken up by the colonic epithelium where it is actively metabolised to produce energy(3,4). Much attention has been devoted to the anticancerous properties of butyrate in human nutrition despite numerous papers suggesting its role as a growth factor. The effects of butyrate, however, depend on the experimental model (in vivo, invitro), the state of the cells (normal or cancerous)(5), the degree of inflammation(6) and the doses used(7). In addition to the recognised effects on intestinal metabolism, butyrate shows indirect effects that contribute to the general metabolism of animals. In all tissues butyrate is a natural component of cellular metabolism. It may also act as a growth promoter when added to diets at low doses (0·1–0·5 g/kg)(8). The purpose of the present paper is to give an overview of the properties of butyrate either originating from bacterial intestinal fermentation or added to the rations of animals. In addition to the recognised effects on intestinal metabolism, butyrate shows indirect effects that contribute to the general metabolism of animals. Production and delivery of butyrate in the gut Butyrate production patterns in the gut at birth and during development In utero, the intestine of the mammalian fetus is sterile. At the time of delivery, the intestinal microbiota are acquired by swallowing maternal anal or vaginal organisms(9). In poultry, hatching through the eggshell has a similar effect. The effect of the first inoculum may be lifelong, directing the development of the immune system and the normal intestinal microbiota. The bacterial composition of the inoculum received at birth, the structure of the host’s intestinal epithelium and the diet all affect the density and composition of the microbiota(10). These microbiota, particularly in the large intestine, have a fundamental role in supplying energy to the host through anaerobic fermentative processes producing SCFA(11). Another source of SCFA in general and butyrate in particular is the milk. Butyric acid is a natural component of mammalian milk, except for human and sows’ milk where only traces can be found. In cows’ milk butyrate concentration is about 0·16 g/l(12), which seems to be insufficient to cover the energy needs of the GI epithelium in the newborn calf but it may be enough to stimulate GI development together with milk-borne hormones and growth factors such as insulin, leptin, glucagon-like peptide (GLP)-2, insulin-like growth factors, epithelial growth factor and many others(13). Another source of butyrate in suckling neonates is the hydrolysis of milk lipids by salivary and gastric lipases. This source of butyrate is difficult to estimate but it may be utilised in the upper GIT. In newborn rats, very high levels of butyrate b-oxidation are detected in colonic epithelial cells. The high oxidation levels may be explained by the fact that the animal is geared to use luminal substrates (high-fat milk diet) with maximum efficiency(14). The stools of normal full-term infants contain less than 100 mM-SCFA(15), with acetate being the major and butyrate the minor acid found in the faeces of formula fed-babies, while virtually no butyrate is found in the faeces of breast-fed infants(16). In premature infants deficient in intestinal lactase, high concentrations of SCFA are produced due to carbohydrate malabsorption, bacterial overgrowth and poor intestinal motility and can significantly lower the pH of the intestinal luminal contents. When specific pathogens, such as Clostridium difficile, proliferate in this environment, they can cause necrotising enterocolitis(17). In healthy infants, fermentation is lower than in adults and butyrate production is established at a slower rate than for acetate and propionate but, by 2 years, an adult SCFA profile has emerged(18). Similar SCFA production patterns have been detected in animals. In broilers aged 1 d, SCFA increased from undetectable levels to high concentrations in broilers aged 15 d, and then stabilised(19). Butyrate production in the gut of adult animals Large bowel fermentation: the main source of available butyrate. In ruminants, the resident microbiota components are essential, as they convert the feed components into nutrients that are readily available for the animal. In single-stomached animals, less microbial involvement is needed, as most nutrients in the diet are directly available to the host. Whilst bacteria affect the performance of the host by competing for dietary compounds, they also provide energy to the host through fermentation. Based on the location of fermentation, animals can be classified into forestomach fermenters and hindgut fermenters. Forestomach fermenters such as ruminants, camels, hippopotamuses and some monkeys have a fermentation chamber cranial to the acid-secreting part of the stomach. Rodents, horses, elephants and most omnivores and carnivores are hindgut and large intestine and/or caecum fermenters, with the GIT serving as a chamber for microbial action and SCFA production(1). Elsden et al. (20), 60 years ago, compared total SCFA concentration at different sites in the GIT of a number of herbivorous and omnivorous species and noticed the highest concentrations in the caecum, and a progressive decline along the colon. In mice and rats, a higher concentration of SCFA was reported in the caecum compared with the colon(21). While in ruminants the highest concentration of SCFA was found in the rumen, another peak was measured in the caecum and colon, indicating that even in forestomach fermenters considerable further digestion occurs in the large intestine(1). The intestine contains several different bacterial species. Saccharolytic bacteria, leading to linear SCFA, CO2 and H2, predominate in the proximal colon, while the majority of proteolytic bacteria, yielding branched SCFA, CO2, CH4, H2, phenols and amines, are mainly in the distal colon where fermentable carbohydrates are depleted on transit(22,23). Polysaccharides, oligosaccharides and disaccharides resistant to the action of hydrolytic enzymes in the upper GIT enter into the caeco-colon. Here, the carbohydrates aredepolymerised to their constituent sugars by a wide range of bacterial cell-associated (Fig. 1) and secreted hydrolytic enzymes such as b-fructosidase, b-galactosidase, xylanase and other hydrolases, and are then fermented(24). Energy is gained in the form of ATP by substrate-level phosphorylation during oxidative substrate breakdown via the fermentation process. The transfer of the resulting reducing equivalents into the metabolic intermediates leads to the formation of large amounts of reduced endproducts such as butyrate, which acts as a terminal electron acceptor in an environment limited in O2. There are two distinct pathways for the production of intracellular butyrate in bacteria. In the first pathway butyryl-CoA is converted to butyrate with the intermediate formation of butyryl phosphate by the enzymes phosphotransbutyrylase and butyrate kinase(25). In the second pathway the enzyme butyryl-CoA:acetate CoA transferase is involved and transfers the CoA moiety from butyryl-CoA to external acetate, which leads to the formation of acetyl-CoA and butyrate. The CoA transferase route is shown to be more dominant in the human butyrateproducing microbiota(26). In vitro data show that strains possessing the CoA transferase convert 75 % of the supplied glucose into lactate when acetate is absent. In the presence of acetate, mainly butyrate is produced(27). Substrates used for bacterial fermentation with butyrate as endproduct. In the large intestine, the ubiquitous nutrient glucose is only of secondary importance. Enterocytes use both glutamine and glucose as the favoured metabolic fuels, while colonocytes preferentially use butyrate(4). The rate and amount of butyrate produced along the colonic lumen depends on the microbiota composition, the chemical composition, the physical form and the amount of substrates available in the diet. Substrates can be indigestible or digestible carbohydrates and, additionally, sloughed cells, mucus and endogenous secretions may also provide some fermentable substances. Examples of indigestible carbohydrates are oligosaccharides (for example, raffinose, oligofructose, inulin) and NSP which can be soluble (for example, -glucans) or insoluble (for example, cellulose and hemicellulose). The former are highly fermentable and hence generate greater quantities of SCFA in the colon while the latter have a rather low fermentability but increase the faecal bulking and decrease the colonic transit time. The most important digestible carbohydrate is starch, known as resistant starch (RS; D-glucose units connected by a-1,4/a-1,6 glucosidic bonds) when it has escaped digestion and/or absorption in the small intestine. RS can be subdivided into four types: physically trapped starch (in coarse grains), RS granules naturally rich in amylose (i.e. raw potato flour), retrograded starch (i.e. cooked and cooled potato) and chemically modified starch (i.e. processed foods)(28). As a result of increasing concentrations of acidic fermentation products, the luminal pH in the proximal colon is lower. This pH seems to boost the formation of butyrate, as mildly acidic pH values allow butyrateproducing bacteria to compete against Gram-negative carbohydrate-utilising bacteria such as Bacteroides spp.(29). An in vitro fermentation study, using faecal inoculum, showed that the population of butyrate producers, as well as the concentration of butyrate, was significantly higher at pH 5·5 while Bacteroidetes dominated as the fermenter was at pH 6·5. Lowering the pH value further, for example, to pH 5·2, resulted in the loss of lactateutilising bacteria, such as Eubacterium hallii, leading to the accumulation of lactic acid(29). Although all dietary fermentable carbohydrates reaching the hindgut have the potential to be converted to butyrate, not all are equally butyrogenic. In vitro (30), animal(31) and human(32) studies have shown that RS fermentation generates relatively more butyrate than NSP fermentation. Butyrogenic substrates may affect the fermentative metabolism in individual bacteria, as more substrate leads to enhanced butyrate formation, which provides a route for the disposal of excess reducing equivalents (i.e. butyrate is used as a hydrogen sink). Butyrogenic substrates may also affect the microbial population by stimulating butyrate-producing species that are efficient primary degraders of these substrates(33). Indirect stimulation of butyrate production occurs by metabolic cross-feeding. The first mechanism is via crossfeeding of oligosaccharide breakdown products released by active polysaccharide-metabolising bacteria. These breakdown products are then fermented by butyrate producers. This was demonstrated in in vitro cultures between bifidobacteria and butyrate-producing species(34). The second mechanism is via cross-feeding of fermentation products such as lactate, ethanol and succinate. These compounds are intermediates in the global fermentation process in the microbiota and are detected in very low concentrations in healthy subjects. In vitro, the accumulation of lactate is prevented when human butyrate-producing strains such as Eubacterium hallii and Anaerostipes caccae are grown in co-culture with lactate-producing strains such as Bifidobacterium adolescentis (35). Scientific reports support the view that RS is the most powerful butyrogenic substrate(36). Both in vitro as well as in vivo fermentation of RS generally results in butyrate production in the order of 20 to 28 mol% compared with about 10 to 15 mol% for NSP(37). In rats fed RS-containing diets, there was a significant increase of the number of bifidobacteria and lactobacilli. Although these bacteria are predominantly acetate and lactate producers and not butyrate producers, butyrate production was significantly enhanced. Therefore it was accepted that the increased number of lactic acid bacteria induced a higher production of lactate that was efficiently converted to butyrate by lactic acid-utilising bacteria(38). Due to the variable structure and complexity of starch, the exact starch type has a considerable impact(37). Among RS, the retrograded RS (RS3) seems to be the most powerful butyrogenic substrate(37). The butyrogenic properties of RS3 is linked to the length of the 1,4-a-D-glucans chain. Chain lengths with a degree of polymerisation of about 20–35 units of glucose are optimal for a high output of butyrogenic RS3(39). It is possible to increase the butyrate production from RS3 fermentation by an appropriate hydrothermal treatment. In a recent study, Jacobasch et al. (39) compared the physiological effects of three RS3 in rats: Novelose 330, annealing-Novelose and heat moisturetreated Novelose. In the caecum, proximal colon and distal colon treated with RS3, the digestive concentration of butyrate significantly increased and in the distal colon, heat moisture-treated Novelose yielded the highest butyrate concentration. This result is of great importance because a high butyrate concentration in the distal part of the colon is considered to be essential for colonocyte health and may have many implications in the prevention and the treatment of some chronic digestive diseases such as diverticulosis. The ability to produce butyrate from RS is also a dose-related effect and may vary greatly among individuals(40). Oligofructose compounds are also associated with greater butyrate production. Although oligofructose is thought to be selectively fermented by bifidobacteria and, to a lesser extent, by lactobacilli, due to their production of b-fructosidase that cleaves the b-(2-1) bonds present in oligofructose and inulin, stimulation of butyrate-producing bacteria is observed(41). Besides dietary fibres, other substrates using different mechanisms are known to increase butyrate concentration. An example is acarbose, an oligosaccharide that increases the amount of starch entering the colon by acting as an a-glucosidase inhibitor(42). Also, the kinetics of butyrate production varies widely between substrates. For example, fructo-oligosaccharides are known to be rapidly fermented, whereas RS is more slowly degraded and can reach the distal part of the colon(43,44). There is little information to explain why bacterial fermentation of specific carbohydrates, such as RS and oligofructose, selectively increases butyrate production, but it is assumed that interconversion reactions from primarily lactate and acetate are largely involved. In the rumen up to 60 % of butyrate is synthesised directly from extracellular acetate through interconversion reactions(45). Acetate and butyrate are interconvertible, but due to the net gain of ATP, the conversion of butyrate to acetate is more favourable for the microbial metabolism(46). Butyrate-producing microbiota. Culture-independent surveys of the adult human gut microbiota have revealed two bacterial phyla, the Bacteroidetes and the Firmicutes(47), commonly dominating this ecosystem. This is also the case for the gut of at least sixty mammalian species(48). The dominant groups found in the large intestines of human subjects(49), pigs(50) and horses(51) and in the rumen of cattle are the Cytophaga, Flavobacterium, Bacteroides (CFB) group of Gram-negative bacteria, and the Clostridium cluster XIVa (Clostridium coccoides) and cluster IV (Clostridium leptum) group of Gram-positive bacteria(10). In poultry, relatives of Clostridium leptum and Clostridium coccoides groups dominate the caecum(52). A high frequency of the Sporomusa group (Clostridium cluster IX), which includes major propionate producers, was reported while no numbers of the CFB group were detected in the avian caecum(53). Sonnenburg et al. (54) demonstrated that members of the Bacteroidetes phylum harbour a large set of genes which encode functions necessary to detect, bind, degrade and import carbohydrates encountered in the gut habitat – either from the diet or from host glycans associated with mucus and the surfaces of epithelial cells. The substrate breakdown products released by those primary degraders may be used by a myriad of other bacterial groups including the butyrate-producer group. The butyrate-producing bacteria cultured so far are strictly anaerobic firmicute bacteria, generally regarded as difficult to grow in vitro. The bacteria are Gram-positive, having a low mol% guaninecytosine (G-C) content and are widely distributed across several clusters within the order Clostridiales. Those clostridial clusters (I–XIX) form a new nomenclature that rearranges the grouping of bacteria based on studies of the evolutionary relationships by 16S rRNA sequencing(55). The bulk of the butyrate-producers belong to cluster IV and cluster XIVa and include some potentially important butyrate producers related to Faecalibacterium prausnitzii, Eubacterium rectale and Roseburia species, respectively(56). In humans, the butyrate-producing bacteria related to F. prausnitzii comprise 5–15 % of the total microbiota(49) and 5–10 % of the total microbiota species are related to Eubacterium rectale and Roseburia (57). In recent studies of poultry, two butyrate producers were described: Butyricicoccus pullicaecorum(58), related to F. prausnitzii, and A. butyraticus (59), closely related to A. caccae, the lactate-utilising bacteria routinely detected in faecal samples from healthy individuals(10). In the rumen of cattle and sheep, predominating butyrateproducing bacteria belong to cluster XIVa and are represented by Butyrivibrio fibrisolvens strains(60,61). Butyrivibrio fibrisolvens is also present in the faecal flora of man, rabbits and horses(62). Depending on the environmental conditions, fermentative metabolism of butyrate producers can result in butyrate, formate, H2, CO2, while lactate and acetate can be either produced or consumed(56). Butyrate producers are considered to play an important role in maintaining gut health mainly through the production of butyrate. Other traits beneficial for the hosts’ health are their capacity to stabilise the luminal pH by consumption of lactate or the anti-inflammatory effect of some butyrate producers(63). Given the importance of butyrate to colonic epithelial metabolism, it is reasonable to assume that butyrate-producing bacteria are well tolerated by the innate immune system(64).
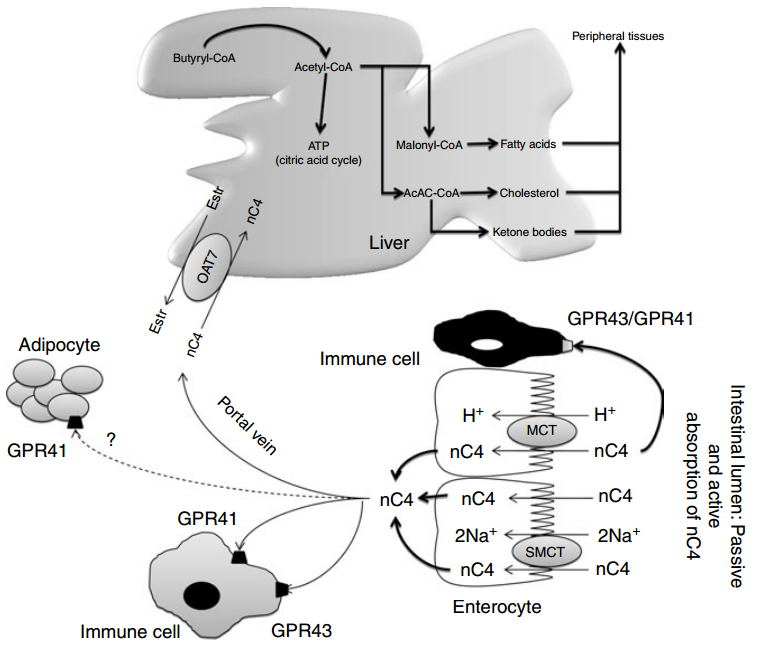
Absorption, transport and metabolism of butyrate Absorption of butyrate, SCFA membrane receptor and transport proteins Earlier studies have demonstrated that SCFA are absorbed in both the small and large intestine by similar mechanisms(65,66). More recent studies suggest the existence of species differences and different transporter isoforms expressed in enterocytes along the intestine(67). On the apical membrane of the epithelial cell, distinct transport mechanisms have been reported, i.e. non-ionic diffusion and mechanisms involving SCFA/HCO3 2 exchangers, monocarboxylate transporter (MCT) type 1(68,69), and Na-coupled MCT(70) (SMCT or SLC5A8/12). On the basolateral membrane, a carrier-mediated, HCO3 2-gradient-dependent anion–butyrate exchange system (Fig. 2) is found(71). The human intestine constitutes MCT3, MCT4 and MCT5 isoforms (the MCT6 expression was not found), with low expression of MCT3 in the ileum, and high expression of MCT4 and MCT5 found predominantly in distal colon(67). In ruminant species, the SCFA-transporting mechanisms in the rumen epithelium are highly efficient, and lead to absorption of nearly all volatile fatty acids. Anion competition experiments in the washed ovine reticulorumen segments revealed that SCFA can be transported by both bicarbonate-dependent and bicarbonate-independent protein-coupled mechanisms(72). The latter was not coupled with MCT. MCT are involved in butyrate transport in pig and human colonic luminal membrane. Ritzhaupt et al. (68,69) demonstrated that butyrate uptake is via a pH-activated, electroneutral anion exchange system. The optimal pH for the activity of the colonic butyrate transporter seems to be 5·5. Butyrate transport with MCT is saturable, coupled with Hþ and inhibited by several monocarboxylates such as acetate, propionate, pyruvate, L-lactate and a-ketobutyrate. More recently, a second class of MCT was identified(70), named SMCT. Two proteins have been cloned: SLC5A8 or SMCT1 and SLC5A12 or SMCT2(73). Conversely to MCT, SMCT transport mechanism involves Naþ uptake by the transport cycle and also uses nicotinate and ketone bodies as substrates. In the colon, Li et al. (17) showed that SLC5A12 (or SMCT2) functions as a tumour suppressor. Ganapathy et al.(70) demonstrated that a non-malignant colon cell line expresses the transporter contrary to malignant cells. So, exposure of non-malignant cells to butyrate does not induce apoptosis. However, when SLC5A12 (SMCT2) is ectopically expressed in malignant cells and when butyrate is added in the culture medium, cells undergo massive apoptosis. In the normal colon, SLC5A8 is claimed to have less importance in butyrate transport than MCT1(74) but it plays a key function in intestinal lactate absorption. In humans, SLC5A8 is considered a tumour suppressor(75). During the transformation of non-malignant cells into malignant ones, expression of SLC5A8 is silenced perhaps to avoid butyrate inhibition ATP (citric acid cycle) Butyryl-CoA Acetyl-CoA Malonyl-CoA Fatty acids Cholesterol Ketone bodies AcAC-CoA Peripheral tissues Liver Estr Estr nC4 nC4 Adipocyte Immune cell GPR41 Portal vein Immune cell GPR41 GPR43 Enterocyte GPR43/GPR41 OAT7 nC4 nC4 nC4 nC4 H+ nC4 nC4 nC4 H+ 2Na+ 2Na+ MCT SMCT Intestinal lumen: Passive and active absorption of nC4 ? Fig. 2. Absorption of n-butyrate (nC4) in the large intestine and subsequent metabolism. Butyrate transport with monocarboxylate transporters (MCT) is saturable and coupled with Hþ transport. Several receptors for butyrate have been identified and detected in a variety of tissues including fat tissue, but the highest expression has been found in immune cells. Butyrate prevents obesity and decreases body fat mass in mice, but the exact mechanism is unknown. After the intestine, butyrate can be metabolised in the liver to produce fatty acids, cholesterol and ketone bodies. In the peripheral blood, no significant concentration of butyrate is found. Oestr, oestrone sulfate; OAT7, organic anion transporter 7; GPR, orphan G protein-coupled receptor for SCFA; SMCT, Na-coupled monocarboxylate transporter. Adapted from Gupta et al. (73)
of histone deacetylase. But in the presence of butyrate, expression of SLC5A8 is again enhanced and it increases cell apoptosis(75). Nevertheless, the lack of susceptibility for colon cancer in SLC5A8 2 /2 mice and the failure to detect a significant uptake of butyrate by SLC5A8 question the role of SLC5A8 both in butyrate transport and colon cancer suppression(76). Two orphan G protein-coupled receptors (GPR) for SCFA, GPR41 (or FFA3) and GPR43 (or FFA2) have been isolated from the intestine(77 – 82). FFA2 mRNA was detected in a variety of tissues, but the highest expression was found in immune cells, including polymorphonuclear cells, suggesting that SCFA might be involved in the activation of leucocytes. Indeed, recent studies in (FFA2) GPR43- deficient mice (Gpr432/2) indicated that acetate administration may help in resolution of the inflammatory response(83). FFA3 has an even more widespread expression pattern than FFA2, including adipose tissues, pancreas, spleen, lymph nodes, bone marrow and peripheral blood mononuclear cells(81). Abundant FFA2 and FFA3 expression was detected in the rat distal ileum and colon, and the human ascending colon(78,84,85). Expression was limited to mucosal cells, and absent in enteric neurons and smooth muscle cells. Immunohistochemical staining with antiFFA2 and anti-FFA3 sera showed FFA2 and FFA3 immunoreactivity in enterocytes and enteroendocrine cells of open type(78,86). The FFA3 immunoreactive enteroendocrine cells were less numerous than the FFA2 cells, and double-immunostaining for FFA2 and FFA3 revealed no common localisation in one cell. Both the FFA2- and FFA3- immunoreactive enteroendocrine cells exhibited peptide YY, but no 5-hydroxytryptamine, expression, supporting the evidence for stimulation of peptide YY release by SCFA(78). The FFA2 and FFA3 distribution and physiological role in the GIT, involving sensing of luminal content, intestinal motility, secretion and innate immunity, have been recently reviewed by Karaki & Kuwahara(86). In healthy rats fed a fibre-free or a RS-enriched diet and using [1-13C]butyrate intra-caecal perfusion, the flux of butyrate production was 0·7–1·8mmol/min whereas portal flux was from 0·2 to 0·4 mmol/min and parietal utilisation from 0·9 to 1·8mmol/min(87). In unfed pigs with an intracaecal perfusion of [1-13]C-butyrate, the parietal utilisation of butyrate varied from 60 to 120 mmol/min when the concentration of perfused [1-13]C-butyrate varied from 0 to 160 mmol/min (L Martin, unpublished results). Energy source for colonocytes In vitro studies have demonstrated that butyrate also represents the preferred energy-providing substrate for the colonic cells(4,88). Colonocytes exhibit a great capacity to rapidly metabolise butyrate. Through fatty acid oxidation, butyrate is entirely oxidised into CO2 or used as a precursor for lipid synthesis(89). In fact, butyrate is able to increase lipogenesis from acetyl-CoA or ketone bodies synthesis via the hydroxyl-methyl-glutaryl-CoA pathway(90). Consequently, the synthesis of many key components of the intestinal epithelial tissue depends on butyrate metabolism (see later). As butyrate is a key substrate for colonocytes, very small quantities reach the general circulation or portal vein. Guilloteau et al. (8) did not observe any changes in plasma butyrate concentration in the peripheral circulation in calves. Nevertheless, the concentration of butyrate in the portal vein may vary according to the diet. Some substrates such as RS or oligosaccharides lead to substantial increases in butyrate concentration in the portal vein(91,92). Hepatic uptake of butyrate is almost total. In the liver, butyrate metabolism also yields acetyl-CoA as in colonocytes. In contrast to single-stomached animals, in vivo studies in steers revealed important butyrate uptake in the rumen as well as limited capacity to metabolise butyrate in the ruminal epithelium and liver. In cattle, butyrate absorption is saturable and if it exceeds the metabolic capacity, it affects rumen epithelial, hepatic nutrient metabolism and the nutrient supply of peripheral tissues(93). Butyrate transport into the peripheral blood circulation: is there proof for physiological direct effects? As shown by Kristensen & Harmon(93) in cattle, substantial amounts of butyrate produced by rumen microbiota may be absorbed through the ruminal epithelium and induce a number of effects via the blood circulation. First, included at a low dose in the diet (0·3 % of DM), butyrate disappears from the upper GIT (mainly in the stomach). It is thought to be metabolised in the GIT wall because it is not found in blood(8,94). Nevertheless, when ingested with the diet, in specific conditions, butyrate would be measurable in peripheral blood(95) and seems to be able to act on peripheral organs (skeletal muscle, brown adipose tissue, liver, etc). Second, butyrate generated from dietary fibre fermentation at a high dose in the hindgut lumen (from 3 to 70 mM) (96) is quickly absorbed in rodents(97) and transported via the portal vein to the liver. In humans the butyrate concentration in the portal vein is about 30 mM whereas it is 12 mM in the hepatic vein(1). A significant amount of butyrate is detected in the portal vein but not in the peripheral circulation in pigs fed a diet enriched with resistant potato starch(98). Similarly, using rye fibres as a RS source, butyrate concentration significantly increases in both the portal vein and peripheral circulation(99). In pigs, the direct perfusion of sodium butyrate in the caecum induces a dose-related increase of butyrate in the portal blood (L Martin, unpublished results). Butyrate, however, is not regularly detected in the peripheral blood even after a long period of perfusion with a supra-physiological solution of butyrate in the colonic lumen (L Martin, unpublished results). Taken together, these data suggest that butyrate can be absorbed from the gut and entirely metabolised either in the gut mucosa or in the liver, which makes a direct effect of butyrate via blood circulation unlikely. Indeed, Bloemen et al. (100) showed that in human patients, no intestinally produced butyrate escaped the splanchnic area due to a highly efficient hepatic uptake. Butyrate and hepatic metabolism Only a small fraction of luminal butyrate can reach the liver via the portal vein. Nevertheless, the effect of butyrate on hepatic cell has been studied. In the liver, butyrate leads to The multiple effects of butyrate 371 Nutrition Research Reviews https:/www.cambridge.org/core/terms. https://doi.org/10.1017/S0954422410000247 Downloaded from https:/www.cambridge.org/core. IP address: 113.52.96.90, on 30 Apr 2017 at 03:08:21, subject to the Cambridge Core terms of use, available at the production of acetyl-CoA that enters into the citric acid cycle. It has been shown that the production of acetyl-CoA via the butyryl-CoA synthase pathway consumes 1 ATP/mol but due to the reoxidation of reduced compounds, the tissue ATP content is maintained. In an experiment using isolated perfused liver of rats, Beauvieux et al. (101) observed that, unexpectedly, butyrate perfusion led to an impairment in energy metabolism, i.e. a decrease in net ATP content in comparison with acetate. The authors hypothesised that butyrate might impair mitochondrial activity inducing an uncoupling between the respiration chain and ATP synthesis. So, the effect of butyrate on hepatic metabolism appears to be close to that of longer-chain fatty acids. As long-chain fatty acids play a part in the pathogenesis of insulin resistance, the effect of butyrate should be considered in specific nutritional conditions (acarbose treatment, high level of dietary RS, etc)(102). When ingested, butyrate enhances glycogen synthesis in the liver at the same rate as glucose(103). The authors demonstrated a clear effect on both a decrease in glucose oxidation and an increase in hepatic glycogen storage. They postulated that such a mechanism could be one of the molecular bases to explain the effect of dietary fibres on the prevention of insulin resistance. Recently a new organic anion transporter (OAT7) was specifically identified in the liver(104), exhibiting a significant transport activity for butyrate (Fig. 2). This transporter mediates the bidirectional transport of oestrone sulfate in exchange for butyrate. This exchange with oestrone sulfate might suggest a contribution (direct or indirect) of butyrate in liver steroid hormone metabolism. More fundamentally, the narrow substrate selectivity of OAT7 suggests that butyrate might participate in the efficient translocation of some sulfate conjugates without interference from the other anionic compounds such as bile salts. In other words, butyrate may play a role in the efficiency of liver detoxification pathways. Effects of butyrate on gastrointestinal epithelial cell functions Several studies have indicated that butyrate, besides providing epithelial cells with energy, markedly increases epithelial cell proliferation and differentiation, and improves colonic barrier function(105 – 108). Butyrate operates as a signal metabolite in the homeostasis of colonocytes, regulating the balance between proliferation, differentiation and apoptosis(109) (Fig. 3). Cell proliferation Earlier studies suggested that trophic effects on the GI epithelium are a result of SCFA. Butyrate is considered to have the strongest effect, that of acetate is weaker, and that of propionate is the weakest(110). Butyrate ingestion is known to modify the microstructure of the small and large intestines in animals and humans(111 – 115). In enterocytes. In the small intestine, butyrate enhances proliferation, differentiation and maturation, and reduces apoptosis of normal enterocytes, mediated through its influence on gene expression and protein synthesis(116). In the small intestine, the crypt depth and the villi height increase largely in butyrate-supplemented weaning or weaned pigs(94,117). Interestingly, when infused in the colon, butyrate was shown to exert trophic effect on ileal and jejunal epithelial cells, presumably indirectly, through a neurohormonal mechanism(114,115). More recent molecular studies investigating cell proliferation, damage, and programmed death (mainly apoptosis) revealed that butyrate speeds up intestinal mucosa maturation during the development or repair after injury(112). Calves supplemented with low doses of sodium butyrate showed a significantly increased mitotic index in the upper jejunum but not in the duodenum or ileum as compared with controls(8). However, changes in the mitotic index were not reflected in villi length and tunica mucosa thickness. In colonocytes. In contrast to the upper intestine, in the large intestine the trophic effects appeared only at the place of butyrate administration. In rodents and swine, all studies showed that the mass of large-bowel tissue is higher in animals fed a butyrate-producing diet, compared with controls(118 – 120). An increase in the length of the colon is repeatedly reported. Increased butyrate concentrations are associated with morphological changes of the mucosa(121). The crypt depth is significantly increased in pigs fed a raw potato starch diet associated with a decrease in the apoptotic activity. The secretory activity, but not the number, of epithelial growth factor-secreting goblet cells is nearly 2-fold increased compared with the controls. Simultaneously, the activity of two anti-apoptotic-regulating proteins is increased (Bcl-2; Bak). An overall reduction of apoptosis was detected without any effects on mitosis. Therefore, it was concluded that the main butyrate effect, in non-tumoural cells, is a shift of apoptosis towards the middle and basal compartments of the crypts. Conversely, in the absence of butyrate, there is massive apoptosis of colonocytes(122). Two plausible mechanisms are used to explain the increased proliferation index of colonic mucosa in physiological conditions(123). The first mechanism is that butyrate is the main energy source for colonocytes and more energy stimulates cell growth. The second mechanism involves the stimulation of the release of GI peptides and/or growth factors by butyrate acting on cell proliferation. Apoptosis In normal cells, butyrate acts as a survival factor but it is an inducer of apoptosis in human colonic carcinoma cells(122). In an in vivo study in pigs, inulin-coated butyrate led to a reduction of apoptosis in the ileal mucosa and an increase in the length of ileal villi although it had no effect on cell mitosis(118). Bailo´n et al. (124) evaluated the effects of butyrate on the proliferation and activation state of different cell types involved in inflammatory bowel disease: intestinal epithelial cells, macrophages and T-lymphocytes, using primary non-transformed cultures and established cell lines. Low concentrations of butyrate inhibited the proliferation of all immune cell types, whereas it induced apoptosis only in activated T-lymphocytes, non-differentiated epithelial cells and macrophage cell lines, but not in differentiated epithelial cells or primary macrophages. The induction of apoptosis was mediated by caspase-3/7 activation (mitochondrial pathway). Butyrate was only able to modify cell activation, measured as expression of inflammatory cytokines, in those cell types in which apoptosis was induced. Apoptosis is an important regulatory process against cancer but various mechanisms can lead cells into a proapoptotic state. Zeng & Briske-Anderson(125) showed that prolonged butyrate treatment in vitro (HT1080 cells) increased the expression of both pro-metastatic and antimetastatic genes but the net effect was an inhibition of pro-metastatic and an activation of anti-metastatic genes. Butyrate also arrested cell growth and migration and invasion of HT1080 cells by inhibiting histone deacetylases, and also by acting on non-histone proteins(126) at the first stages of DNA damage. Once DNA is damaged, butyrate seems to be ineffective. The absolute concentration of butyrate is also very relevant, as low amounts of butyrate enhance cell proliferation while high amounts inhibit it. In the human colon cancer RKO cell line, butyrate induced apoptosis through a mechanism involving the c-Jun-N-terminal kinase (JNK) mitogen-activated protein kinase (MAPK) kinase pathway(127). In contrast, in neutrophils obtained from healthy human volunteers, butyrate increased apoptosis irrespective of their activation state, by factors other than MAPK and GPR, and their mechanisms probably relate to their histone deacetylase inhibition activity, which may control a1 mRNA expression(128). In the large intestine, cell proliferation, differentiation and positional localisation along the crypt axis are governed by several signalling pathways including the Wnt/b-catenin pathway(129). When Wnt signalling is activated, normal cell differentiation and proliferation are modified, leading to probable cancer genesis(129). Furthermore, the relative levels of Wnt signalling determine whether cells proliferate or go to apoptosis(126), since several in vivo studies support the evidence that increased activation of Wnt transcriptional activity enhances apoptosis in some cancerous cells. In vitro studies showed that butyrate hyper-induced Wnt transcriptional activity in some tumour cell lines(130). The authors conclude that both relatively high and relatively low level of canonical Wnt transcriptional activity can lead to cancerous cell apoptosis, and that cells involved in the tumorigenic process due to Wnt signalling are more vulnerable to butyrate action. Beneficial effects on performance and intestinal health Studies with oral administration of butyrate showed that butyrate can improve growth performance in production animals. The effective doses of sodium butyrate were fixed between 1 and 4 % of DM intake, and protection of the molecule by microencapsulation in lipid matrix improved its efficiency. Moreover, slow release of butyrate from a lipid matrix prevented its rapid metabolism in the stomach and upper small intestine, thereby increasing the area under the influence of the molecule(131,132). Butyrate was found to have widespread positive effects on growth, digestibility and feed efficiency through the modulation of cell proliferation, differentiation and function in the GIT, especially mucosal epithelial cells, and on defence systems (barrier function, antimicrobial potency, immune system) in healthy and sick animals(2,94,133 – 138). Effects on growth performance and digestive efficiency. Dietary supplementation with butyrate enhanced growth rate and improved the feed conversion rate in calves before and during weaning(8,131,139) and in piglets(2,140 – 143). Studies in neonatal(112) and in weaned(111,138) piglets showed that the ingestion of butyrate resulted in improved performance, which was further improved if butyrate was fed sooner rather than later post-birth. Butyrate supplementation during the suckling period had no influence on ileal apparent digestibility, but it significantly improved the faecal apparent digestibility of feed components after weaning(111). In these studies, butyrate supplementation delayed gastric emptying, which may suggest that increased gastric retention (longerlasting gastric phase of digestion, slower digesta flow to the intestine) is beneficial for digestive processes(144,145). In the newborn calves, the effect of butyrate supplementation both to the milk replacer and starter diet reduced diarrhoea, and led to an improved health status in the first weeks of life(131). In calves fed milk formula based on soyabean protein, butyrate supplementation improved the DM digestibility and the digestibility of major components of the diet(146). Moreover, the concomitant addition of butyrate to milk replacer and starter mixture may stimulate rumen development. Thus, the weight of the rumen (percentage of the whole stomach) was increased at the expense of the abomasum, and the ruminal papillae length and width were increased(131), improving the absorptive surface of the rumen.